
Biology, 02.03.2020 21:59 btaylor1179
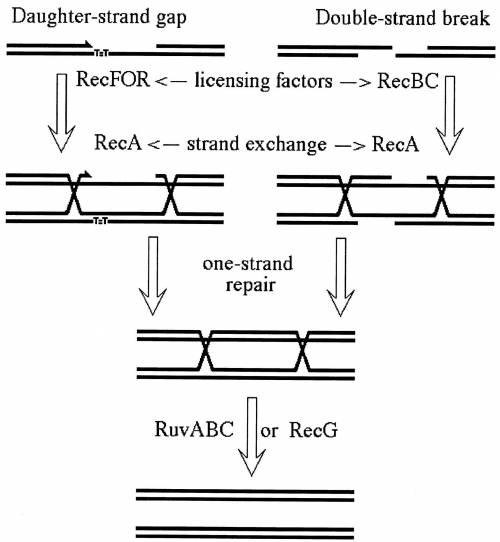
Draw out the double strand break model of recombination, showing the parental and recombinant outcomes using the assignmentprovided in class. What phenotype would occur in cells with the following mutations: recA, RuvA, RuvB, Rec B

Answers: 1


Another question on Biology
Biology, 22.06.2019 11:00
4. if a cell is placed inside a solution that has a higher concentration of solute than on the inside of the cell, what can be said about the movement of water? a. water will move out of the cell, causing it to shrivel. b. water will move in and out of the cell at the same rate. c. water will move out of the cell, causing it to swell. d. water will move into the cell, causing it to swell.
Answers: 2
Biology, 22.06.2019 11:30
In "the pig," what is the main effect that the piglet initially has on kibuka?
Answers: 2
Biology, 22.06.2019 12:30
Which of the following observation darwin shape his concept of descent with modification? a) species diversity declines farther from the equator. b) fewer species live on islands than on the nearest continents. c) birds live on islands located farther from the mainland than the bird's maximum nonstop flight distance. d) south american temperate plants are more similar to the tropical plants of south america than to the temperate plants of europe. e) earthquakes reshape life by causing mass extinctions.
Answers: 1
Biology, 22.06.2019 16:30
During photosynthesis, hydrogen ions are most directly used to in the chloroplast pictured above. a) make glucose b) make chlorophyll c) produce carbon dioxide d) drive the production of atp
Answers: 1
You know the right answer?
Draw out the double strand break model of recombination, showing the parental and recombinant outcom...
Questions

Mathematics, 28.10.2019 20:31

Computers and Technology, 28.10.2019 20:31


English, 28.10.2019 20:31


Mathematics, 28.10.2019 20:31
Mathematics, 28.10.2019 20:31
Mathematics, 28.10.2019 20:31
Social Studies, 28.10.2019 20:31
Mathematics, 28.10.2019 20:31
History, 28.10.2019 20:31


